



|
||||
| Главная | ||||
|
Назад | Оглавление | Вперед | |
| Гостевая | ||||
|
||||
ГЛАВА 4 |
||
| ПОПУЛЯЦИОННО – ГЕНЕТИЧЕСКИЙ АНАЛИЗ МАТЕРИАЛА | ||
|
4.1. Популяционная структура и эффективнорепродуктивный размер Известно, что существенные изменения признаков происходят под влиянием факторов изоляции, миграции, дрейфа генов и естественного отбора. Прежде чем перейти к выяснению механизмов действия этих факторов, необходимо установить временную и пространственную организации армянской популяции, ее эффективный размер, так как эти факторы имеют прямое отношение к воспроизведению генетических особенностей в поколениях. Материалом для характеристики возрастной структуры послужили опросные и демографические данные. Основные данные о возрастной структуре в 15 армянских микропопуляциях приведены в табл. 19 и на рис. 15 (а, б, в, г) для 3 микропопуляций и тотальной армянской популяции. Как видно из таблицы, во всех 15 популяциях наблюдается почти полное совпадение возрастной структуры, что увеличивает надежность данной характеристики популяции. При более дробном делении групп, принятом для построения возрастных пирамид, обнаруживается некоторое различие структуры населения в разных микропопуляциях. Но первое, что бросается в глаза, это то, что подавляющее большинство людей находится в возрасте до 30 – 40 лет. И c наоборот, предельно мал процент людей, находящихся в возрасте выше 55 лет. Это говорит о том, что популяции, исследованные нами, растущего типа. Прямым следствием возрастной структуры является ее средний возраст, который в армянских микропопуляциях равен 27 годам. Он невысок, что можно объяснить высоким темпом рождаемости. Средний возраст популяции должен быть сопоставим с темпом ее реорганизации. Для оценки репродуктивноэффективного размера популяции прежде всего необходимо определить собственно репродуктивную часть ее тотального размера (Рычков, 1974). Данные для определения границ репродуктивного периода приводятся в табл. 20. Во всех человеческих популяциях существует разрыв между физиологическим и социально обусловленным началом репродуктивного периода: максимальная физиологическая нижняя граница репродуктивного возраста у женщин почти совпадает с минимальной социально обусловленной границей, и в среднем этот разрыв для женщин достигает 7 – 8 лет. Велики и половые различия в социально обусловленном начале вступления в репродуктивный период. Поэтому в зависимости от выбора границ репродуктивного периода будут меняться оценки репродуктивной части тотального размера популяций. Мы брали несколько границ репродуктивного периода и проводили оценку репродуктивного объема популяции в смешанных границах. Репродуктивный размер составил в наших популяциях 41% от тотального размера (табл. 21). Надо заметить, что в разных микропопуляциях эта доля колеблется от 23 до 48%. Эффективно-репродуктивный размер армянской популяции оценивался исходя из разброса 4.2. Генные миграции и их структура в Армении Взаимодействие между популяциями возможно благодаря обмену генами – миграциям. В популяционно-генетическом плане взаимодействие мыслится как обмен генами между популяциями, благодаря которому каждая отдельная популяция получает генетически важную информацию о состоянии системы в целом и о состоянии окружающей среды не только в ближайших, но и в крайних, периферических частях ареала. На основе опросных и демографических данных была составлена матрица миграций генов, которая дает возможность количественно оценить интенсивность миграций. Матрица миграций между 36 армянскими популяциями дана в табл. 22 а и б. Каждый пункт охарактеризован коэффициентами прямой и обратной миграций генов, указывающими на степень их изолированности. Соответствующая матрице пространственная структура генных миграций показана на рис. 16, из которого видно, что армянские микропопуляции в действительности связаны в единую систему благодаря обмену генами друг с другом. Но здесь необходимо предположение о типе связи миграций с пространственной структурой популяционной системы. В матрице генных миграций учтены как миграции между конкретными изученными популяциями, так и те, которые зафиксированы в этих популяциях, происходя из других районов Армянской ССР и за ее пределами. Нижние значения в ячейках матрицы характеризуют частоту притока генов в i-тую популяцию, а верхние – частоту отдачи генов из i-той популяции в другие (табл. 226). В данной ситуации, когда числа популяций – источников миграций генов, много больше числа популяций, в которых проводилось исследование, более надежными оценками частоты мигрантных генов являются нижние значения в ячейках табл. 22. В среднем соотношение генов эндогамного (по главной диагонали) и экзогамного происхождения составляет округленно 5:1, а доля генов, принадлежащих по происхождению к конкретной локальной популяции, составляет около 83%. 17%й уровень миградции генов является суммарной оценкой, не скоректированной ни на число популяций – источников этой миграции, ни на степень генетических различий между ними. История образования Армянской ССР и формирования ее населения, сопровождавшаяся значительными передвижениями групп населения, размещением на новых территориях, равно как и значительной миграцией армянского населения из других стран, дает основание исходить в анализе генных миграций из «островного» варианта популяционно-генетических моделей миграционной структуры. Этот вариант предполагает отсутствие зависимости вероятности миграций и величины коэффициента миграций от расстояния, разделяющего популяции. Для такой структуры миграции оценки me могут быть получены по формулам, приведенным ранее (Рычков, Шереметьева, 1974):
Средняя величина коэффициента миграции по совокупности 36 оценок прямых и обратных миграций в 18 изученных популяциях составила Me =.0074, однако следует учесть, что интенсивность прямых миграции на порядок выше, чем обратных. Это означает, что приток генов идет в основном из иных популяций, чем те, в которых изучены признаки дерматоглифики. В связи с этим в табл. 23 коэффициенты прямых миграций в популяции, оцененные, как указывалось выше, более надежно, чем в обратном направлении, подразделы на 2 компоненты. II-ая из компонент скорости прямых миграций связана с обменом генами между популяциями в пределах Армянской ССР и составляет в среднем Другая возможная оценка В работе также использован и «ступенчатый» вариант миграционной структуры Кимуры и Вейса (Kimura, Weiss, 1964). Авторы модели исходят из предположения, что крайне отдаленные популяции не имеют возможности непосредственно обмениваться генами друг с другом, миграции происходят посредством промежуточных популяций и приобретают ступенчатый характер. По миграционной матрице видно, что миграции осуществляются более чем на один шаг. В этом случае авторы рекомендуют пользоваться вариансой генных миграций в качестве меры интенсивности. В нашей работе использован двумерный вариант «ступенчатой» модели.
Для проверки надежности таких параметров популяционной структуры, как Ne и Me, мы сравниваем эмпирическую вариансу частоты генов с теоретической предполагаемой вариансой. Варианса, стандартизованная по средней частоте гена – 1) в случае «островной» модели Райта представляет: где по нашим данным
2) в случае двумерной «ступенчатой» модели Кимуры и Вейса Где,
где К означает связанный эллиптический интеграл первого порядка.
Величина Надо учитывать то обстоятельство, что предсказывая теоретические вариаисы «островного» и «ступенчатого» типов структуры армянской популяции, мы исходили из эмпирических демографических данных о размере и генных миграциях популяции. Ожидаемый уровень генетической дифференциации можно определить, используя частоту фамилий в популяции (Рычков и соавт., 1974, 1976), т. к. Получаемая где xk – средняя частота k-той фамилии в тотальной популяции, В нашем материале
где n(к) – число фамилий в тотальной популяции, В нашем анализе учитывались фамилии отцов, так как фамилии матерей не наследуются. Материал представляет 85 фамилий для 20 армянских микропопуляций (см. табл. 24), где новая оценка В таблице 25 представлены результаты оценки вариабельности частот изученных признаков по локальным популяциям Армении в виде 95%го доверительного интервала для средней частот Таким образом, уровень межпопуляционной изменчивости населения Армении оказывается одним и тем же при двух разных системах данных, используемых для его оценки – дерматоглифических и фамильных. Средняя из 85 оценок стандартизованной вариан сы частоты фамилий Чтобы понять происхождение этой изменчивости, необходимо построить такую модель генетического процесса населения Армении, которая позволила бы предсказать ожидаемый уровень дифференциации населения при выполнении посылок, заложенных в эту модель. Совпадение оценок стандартизованных варианс частот дерматоглифических признаков и частот фамилий указывает на то, что источник наблюдаемой изменчивости не связан специфически ни с одной из этих систем признаков и, скорее всего, заключается в определенном соотношении двух противоположно направленных процессе – случайного дрейфа генов в локальных популяциях Армении и обмена генами между ними. Исходя из соотношения двух параметров популяционной структуры – генетически эффективного размера Ne и скорости миграций генов Me между ними, в табл. 26 представлены теоретически ожидаемые значения стандартизованной генетической вариансы при «островном» и «ступенчатом» вариантах миграционной структуры. Здесь же приведены эмпирические оценки стандартизованных, варианс по признакам дерматоглифики Таким образом, наблюдается хорошее соответствие эмпирически зафиксированных значений F статистик для двух различных систем наследуемых признаков с теоретически ожидаемыми значениями варианс при «островном» типе структуры и достоверное отличие при «ступенчатом» типе структуры. Рассматривая среднюю из варианс по большому числу признаков, мы как бы избавляемся от действия отбора. Это совпадение касается среднего значения На заключительном этапе нашей работы мы попытались реконструировать время для достижения существующего ныне уровня дифференциации армянской популяции по признакам дерматоглифики. Если эти признаки наследуются без отклонений, значит, они дошли до нас из глубин веков, и было бы интересно узнать, когда сформировался переднеазиатский комплекс дерматоглифических признаков и какое время потребовалось для возникновения различий этих признаков, которые присущи современным микропопуляциям армян. Воспользовавшись методами, изложенными ранее (Rychkov, Sheremetyeva, 1977), и используя результаты варианс генных частот, мы подсчитали время достижения нынешнего уровня дифференциации армянской популяции по признакам дерматоглифики. Известно, что темп изменения во времени зависит от эффективнорепродуктивного размера Ne. Для этой цели мы подсчитали величину эффективного размера в выборках армянских микропопуляций, исследованных по дерматоглпфнческим признакам (табл. 29). Эта величина в среднем составила 49 человек, т. е. опять таки 31% от тотального размера выборки, как и в случае доли репродуктивно-эффективного размера армянской популяции в целом от тотального размера населения по данным демографии. Величина Ne=49 в выборках достаточно мала, что благоприятствует случайному дрейфу генов, которому, в свою очередь, могут противодействовать генные миграции, связанные с особенностями популяционной структуры армянского народа, в которой интенсивные миграционные процессы стали основой формирования населения. Располагая генетической информацией об уровне различий дерматоглифических признаков, наследовании фамилий и миграционной структуре, процесс внутренней дифференциации армянских микропопуляций можно выразить следующей формулой:
где t – темп смены поколений, т. е. число поколений, донесших до нас все генетическое разнообразие, накопленное за сотни и тысячи лет. При эмпирической стандартизованной вариансе по признакам дерматоглифики в случае фамильной структуры темп смены поколения – t = 355.18 лет, Таким образом, можно заключить, что по признакам дерматоглифики и структуре фамилий время дифференциации исследованных субпопуляций от материнской прапопуляции армян насчитывает около 8 – 9 тысяч лет, что позволяет предположить, что уже в неолитическое время на Армянском нагорье был сформирован исходный тип человека, обладающего перед неазиатским комплексом изученных признаков и давший начало современным популяциям армян с присущими им признаками переднеазиатской расы. Можно также предположить, что такой комплекс признаков далее распространился по территории Передней Азии и Кавказа. Такой результат, хоть и хорошо согласуется с данными истории, лингвистики и археологии, все же требует дальнейшей разработки и проверки на других системах антропологического и генетического полиморфизма. |
||
|
||||
| Главная | ||||
|
Назад | Оглавление | Вперед | |
| Гостевая | ||||
|
||||
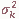 числа гамет относительного к гамет, передаваемых родителям репродуктивного возраста следующему поколению (Рычков и др., 1974). Он отличается от репродуктивоного объема Nr в той мере, в какой существует неравный вклад лиц родительского поколения в генофонд следующего поколения. Это создает разброс
числа гамет относительного к гамет, передаваемых родителям репродуктивного возраста следующему поколению (Рычков и др., 1974). Он отличается от репродуктивоного объема Nr в той мере, в какой существует неравный вклад лиц родительского поколения в генофонд следующего поколения. Это создает разброс 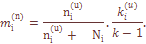
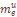 =0.0094. Вторая компонента, составляющая в среднем
=0.0094. Вторая компонента, составляющая в среднем 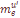 =.0040, характеризует скорость миграции в отдельно взятую популяцию генов переселенцев из Западной Армении. Величина
=.0040, характеризует скорость миграции в отдельно взятую популяцию генов переселенцев из Западной Армении. Величина 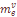 =.0014 характеризует средние значения скорости обратных миграций. Поскольку миграция из-за рубежа не имеет обратного направления, то общий коэффициент миграции генов рассчитывался как
=.0014 характеризует средние значения скорости обратных миграций. Поскольку миграция из-за рубежа не имеет обратного направления, то общий коэффициент миграции генов рассчитывался как 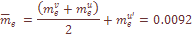
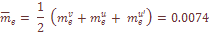
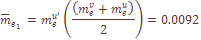
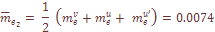
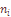 – число миграций в i-тую популяцию или из нее;
– число миграций в i-тую популяцию или из нее;
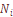 – число людей, происходящих от эндогамных браков;
– число людей, происходящих от эндогамных браков;
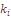 – число реализованных направлений миграции в данную популяцию (u) или из нее (v);
Me – коэффициент миграции в данную популяцию в пределах АрмССР (u), в Арм ССР из Западной Армении (u') или из нее (v).
– число реализованных направлений миграции в данную популяцию (u) или из нее (v);
Me – коэффициент миграции в данную популяцию в пределах АрмССР (u), в Арм ССР из Западной Армении (u') или из нее (v).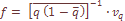
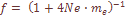 (1)
(1) 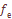 = 0,0577,
= 0,0577, 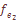 =0,0712;
=0,0712;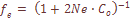 (2)
(2) 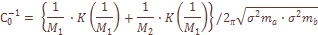
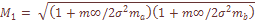
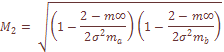
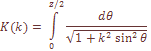
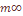 характеризует систематическое давление, испытываемое популяционной системой в целом. При вычислении теоретических варианс давление естественного отбора не принималось во внимание. Мы исходили из гипотезы генетической, дифференциации как селективно-нейтрального процесса, принимая
характеризует систематическое давление, испытываемое популяционной системой в целом. При вычислении теоретических варианс давление естественного отбора не принималось во внимание. Мы исходили из гипотезы генетической, дифференциации как селективно-нейтрального процесса, принимая 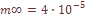 , т.e. несколько выше, чем уровень мутационного давления. Ожидаемое значение вариансы при «островном» типе структуры оказывается равным:
, т.e. несколько выше, чем уровень мутационного давления. Ожидаемое значение вариансы при «островном» типе структуры оказывается равным: 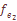 = 0.0712, а в случае “ступенчатого"
= 0.0712, а в случае “ступенчатого" 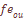 не зависит ни от генетических, ни от демографических данных о размере популяций, ни от предположения о структуре популяций. Фамильная дифференциация популяции является аналогом генетической дифференциации и имеет общую основу с исторически обусловленной структурой популяции и структурой миграции. Оценка Fst через данные о фамильной структуре производилась по формуле:
не зависит ни от генетических, ни от демографических данных о размере популяций, ни от предположения о структуре популяций. Фамильная дифференциация популяции является аналогом генетической дифференциации и имеет общую основу с исторически обусловленной структурой популяции и структурой миграции. Оценка Fst через данные о фамильной структуре производилась по формуле: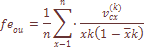
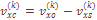 – корректированная варианса частоты k-той фамилии,
– корректированная варианса частоты k-той фамилии, 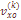 – наблюдаемая варианса;
– наблюдаемая варианса; 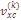 – выборочная варианса, зависящая от метода учета частоты фамилии.
– выборочная варианса, зависящая от метода учета частоты фамилии. 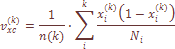
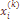 частота k-той фамилии в i-той популяции,
частота k-той фамилии в i-той популяции, 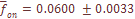
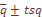 и стандартизированного по частоте
и стандартизированного по частоте 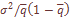 значения систематической вариансы, представляющей разность эмпирического
значения систематической вариансы, представляющей разность эмпирического 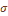 и выборочного значения
и выборочного значения 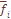 -тая, отметим небольшую ее вариабельность по разным системам признаков, что отражено в малом значении ошибки средней величины (
-тая, отметим небольшую ее вариабельность по разным системам признаков, что отражено в малом значении ошибки средней величины (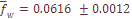 ), составляющей лишь 2% от средневзвешенной по всем системам признаков. Другими словами, уровень дифференциации населения Армении по любой из 8 систем, включающих в общей сложности 35 признаков при 27 независимых, приблизительно один и тот же.
), составляющей лишь 2% от средневзвешенной по всем системам признаков. Другими словами, уровень дифференциации населения Армении по любой из 8 систем, включающих в общей сложности 35 признаков при 27 независимых, приблизительно один и тот же. 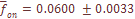 лишь на 2.5% отличается от среднего значения
лишь на 2.5% отличается от среднего значения 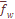 и структуре фамилий
и структуре фамилий 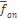 .
. 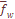 по признакам дерматоглифики. Поэтому необходимо рассмотреть, как отличаются стандартизованные оценки варианс по каждому признаку от установленных теоретических значений дифференциации. Такой анализ эффективен, так как мы можем по хаpарактеру отклонения эмпирической стандартизованной вариансы от теоретических значений стандартизованных варианс судить о подверженности того или иного признака естественному отбору. Как видно из
по признакам дерматоглифики. Поэтому необходимо рассмотреть, как отличаются стандартизованные оценки варианс по каждому признаку от установленных теоретических значений дифференциации. Такой анализ эффективен, так как мы можем по хаpарактеру отклонения эмпирической стандартизованной вариансы от теоретических значений стандартизованных варианс судить о подверженности того или иного признака естественному отбору. Как видно из 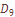 и центральный трирадиус t, находятся под дифференцирующим давлением естественного отбора. Остальные изученные признака – селективно нейтральны. Установленные стабилизирующие и дифференцирующие воздействия, которым подвержены некоторые дерматоглифические признаки, можно рассматривать как проявление фактора генетической адаптации. Необходимо, однако, учесть, что признаки, выделенные как селективно значимые, являются частью соответствующих систем признаков и что в отношении этих систем аналогичный анализ не дает указаний на существенные отклонения от селективнонейтрального уровня дифференциации, предсказываемого значениями
и центральный трирадиус t, находятся под дифференцирующим давлением естественного отбора. Остальные изученные признака – селективно нейтральны. Установленные стабилизирующие и дифференцирующие воздействия, которым подвержены некоторые дерматоглифические признаки, можно рассматривать как проявление фактора генетической адаптации. Необходимо, однако, учесть, что признаки, выделенные как селективно значимые, являются частью соответствующих систем признаков и что в отношении этих систем аналогичный анализ не дает указаний на существенные отклонения от селективнонейтрального уровня дифференциации, предсказываемого значениями 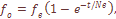
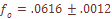 , эффективном размере Ne = 49 и длительности поколения (среднем возрасте) t = 27, время, которое потребовалось для достижения современного уровня дифференциации армянского народа, составит:
, эффективном размере Ne = 49 и длительности поколения (среднем возрасте) t = 27, время, которое потребовалось для достижения современного уровня дифференциации армянского народа, составит: {kind=link}